主题:【原创】死亡起源 The Origin of Death -- az09
家园 非常好! 这两篇的启发性的设问很多是我想过、却没有仔细系统条理地整理分析过的。所以,一看到就被吸引了。谢楼主的工作。
家园 如果是这样,未来人类的合理选择未必是延长生命 如您所说,任何一个物种的寿命是一个平衡的产物:一方面为维持一个物种基本数量,必须有一定的寿命;另一方面为保证物种有稳定、积极进化的趋势,及时淘汰老旧DNA,这个寿命必须有一个上限,不能无限延长。这个动态平衡的结果,决定了具体物种的普遍寿命。
对于人类而言,这个下限比较容易确定--大概40岁左右,即从出生到能够抚育下一代到达生育年龄的最低时限;但上限是多少?什么是老旧DNA,必须予以淘汰以免影响种群质量?
人类的进化已经是以脑力为主,而脑力的进化与DNA的相关性,难以确定 -- 即使是同卵双胞胎,在社会中的发展也会很不一样。而从另外一个角度看,人类发展的最主要推动力量是少数天才,比如爱因斯坦、毛泽东等。过去实践都证明了,天才的出现是随机的 -- 爱因斯坦的儿子绝无可能成为爱因斯坦,天才无法遗传,也就是王侯将相宁有种乎。因此,绝大多数普通人繁衍后代,就是在撒DNA骰子 -- 看自家的儿子是否会是个天才。从这个意义上讲,尽快掷骰子才是最明智的做法 -- 掷的越多,碰到天才的机会越多,也就是加速进化。从生育的角度,就是在力所能及的范围内早生、多生碰运气。但这会迅速造成资源不足,而最佳的解决方案,显然是老一代尽快退出,在生育能力下降前,把资源留给下一代继续碰运气。同时,从个人的角度看,一个人最富有创造力的时候,也就是20-35岁这段年龄,之后的时间,经验或者会继续积累,但创造性很难维持;而把持的资源,不如让给下一代。
从这一点出发,似乎也证明了人类的寿命不应该长于上限太多,或者说人类寿命的下限就是上限,也就是40岁左右。
通宝推:铁手,家园 外祖母的作用 也许下限不应该是40岁,隔代人对下一代的抚养,成长有积极作用,三代人的家庭算是人类完整的家庭。也许下限是60岁?
家园 据此,寿命被重新定义了! 说理非常清晰。不仅阐述了死亡机制本身是一个自杀机制,而且揭示了衰老过程其实就是生物个体的寿命的概念。生物个体寿命的长短不过是一个自杀过程的长短问题。那么,自从人类个体开始衰老,自杀机制就已经被触发了。人类个体寿命之延长,不过是一个具体个体利用其人为创造的种种手段,对抗群体普遍的寿命水平的过程。
但是,群体寿命水平需要通过什么方式来提高?我们也就非常清楚了。因为不同人类群体之间,在物质分配过程中是存在激烈竞争的。而这就导致了阶级的产生,本主题也就进入了社会讨论阶段。
通宝推:acton,老老狐狸,家园 收益不浅啊 兄台确实是大才。
家园 死亡起源(七)—— 迷雾与噪声 续上:死亡起源(六)
3. 衰老和死亡的开关
3.1 迷雾与噪声。
通过分析水螅、水母、线虫这样的低等生物,我们似乎已经找到了衰老和死亡的关键点了。但是,随着生物的演化,生命变得越来越高级,也越来越复杂以后,各种迷雾与噪声就随之产生了。
《Cell》 杂志在2013年5月发表了一篇综述:《The Hallmarks of Aging》(衰老的标志)[14],文中综合叙述衰老和死亡的9个标志。它们分别是:基因组不稳定性,(genomic instability), 端粒缩短(telomere attrition),表观遗传学改变(epigenetic alterations), 蛋白内稳态丧失(loss of proteostasis), 营养感应失调(deregulated nutrient sensing), 线粒体功能异常(mitochondrial dysfunction), 细胞衰老(cellular senescence), 干细胞耗竭(stem cell exhaustion)和细胞间信息交换改变(altered intercellular communication)。相信该文章的作者以及许多人或许就把上面的这些标志作为原因了。不过我们只要细想一下,就会发现,这些标志当中,相当部分可能都只是结果,而不完全是原因。
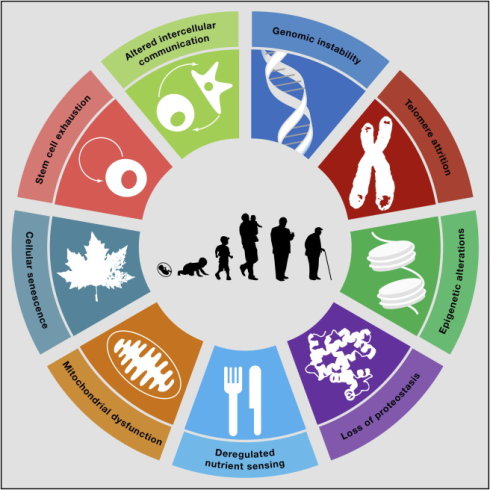 图28. Cell 期刊的文章描述了9种衰老标志/原因
图28. Cell 期刊的文章描述了9种衰老标志/原因 图29. Cell期刊的文章中,针对9种衰老原因的处理措施
图29. Cell期刊的文章中,针对9种衰老原因的处理措施说个题外话。本段一开头所举出的这篇文章《The Hallmarks of Aging》,是世界顶级期刊《Cell》的一篇综述。所谓的综述,就是它不是一篇文章,而是一个集大成者。是这些年无数最顶尖的关于衰老研究的各种文章的集大成者。所有的试图挑战这篇文章的人,首先得扪心自问:自己是不是一个疯子? 即便是俺这个满脑子离经畔道思想的人,也不敢一开始便挑战这篇文章,因为我很清楚,我一定会被人当疯子看待。呵呵。
好在,我们已经从(一)讨论到了(六)了,已经详细讨论了许多关于“永生”、“有机整体的程序化死亡”等等概念了。现在开始质疑这篇文章,我们已经开始有了一些底气了。(另外,这篇综述是2013年5月出的,而我们拿2015年才出来的诸多证据来反驳它,这也多少也有些占便宜了。 呵呵。)
之所以要把篇文章拿出来讨论,是因为这篇文章所叙述的内容,和它所表现出的思想,其影响非常的深远。在承认它的合理性的同时,我们也必须指出它的不足之处。
如果我们只是把眼光局限在细胞内的话,我们或许也可以得出许多可以自圆其说的结论。比如,(六)里面所举的美国西北大学线虫的死亡开关的例子,如果一个研究者,观察到线虫体细胞内的分子变化和蛋白质质量控制的下降,它完全可以就此得出一个简单的结论:是因为细胞内的这些蛋白质的变化,导致了线虫的衰老和死亡。而实际的情况呢,却是真正的原因,是来自于细胞外的,由线虫的生殖干细胞给出的“死亡信号”。
另外,有些细胞死亡的原因,可能已经超出了它的“理想状态"了,这个不在本文讨论的范围之内。总不成拿个探针,把一个细胞破坏掉,然后与我讨论它的衰老与死亡的概念吧。 呵呵。
相信大部分人可能还会坚持,由于高等生物的组织分化的复杂性,生物的衰老和死亡,主要是因为组织结构过于复杂,各种组织、细胞随着时间消耗老化所致。另外,还有人试图用物理学的“熵增大”的概念来解释生命的衰老与死亡。因为根据物理学第二定律,“熵”总是要增大的,物质总是倾向于从有序到无序,所以,象生命这样的复杂系统的崩溃和死亡也就是无法避免的。不过,我认为这种解释是说不通的。因为热力学第二定律的前提条件是一个“孤立系统”。但是,很显然,生命并不是一个孤立系统,与外界的物质和能量交换,乃至新陈代谢,是生命的基本特征和属性。
总之,上面的各种说法,我承认其中有相当的合理性,而且在某种意义上的事实也是如此。但是,事实绝对没有那么简单。实际上生物的衰老和死亡,或许和上述的原因有许多关联,但是起决定性的,还是死亡和衰老机制的适时开启或者关闭。
为了解释这一点,我们可以看一个非常简单的例子:比如人类与狗的比较。人类与狗的基因以及细胞乃至器官结构,有非常多的相同之处。在已识别出的2.4万多个人类基因中,至少有1.8万个与狗的基因相同。从演化的角度看,人和狗拥有共同的祖先,而且两者的亲缘关系还比较近。但是,我们如何解释这样一个现象:一个15岁的小孩,他的全身的细胞和组织器官都处在欣欣向荣的状态,而一只15岁的狗,却已经开始垂垂老矣,器官衰竭,面临死亡?如果简单的以消耗论的观点来看,我们如何解释,结构非常接近的15岁的人类少年的器官,为什么就要比15岁的狗的器官要更年轻,要更有活力?另外,用消耗论的观点,我们也无法解释,为什么15岁的狗的患癌症的概率,要远远超过15岁的年轻人类的患癌症的概率
 图30. 15岁的年轻人,满脸朝气蓬勃
图30. 15岁的年轻人,满脸朝气蓬勃 图31. 一只15岁,面临死亡的老狗,这个年纪的狗有50%都会患有癌症
图31. 一只15岁,面临死亡的老狗,这个年纪的狗有50%都会患有癌症然后,当我们把前面所述的一些衰老标志当作我们身体内细胞和组织衰老和死亡的主要原因的时候,我们不妨再拿出我们身体内的一个细胞来观察。当我们注意到这个体细胞的各种衰老的标志,比如生长缓慢、DNA错误累积、蛋白质出问题、分裂速度变慢甚至不再分裂、细胞凋亡等等,我们却不能忘记一个事实:根据细胞学原理,这个细胞,是从几十亿年前,由某个细胞,一直分裂而来的!它穿过了亿万年的时空,经历过可能超过上百亿次的分裂,以“永生”的方式生存到现在,然后才能被我们观察到。不管这几十亿年当中,有多少它所分裂的细胞死亡,但是它却一直是那个幸运儿,所以它才能活到现在。现在,我们只注意到了它的生命,最终在我们观察下逐渐停止分裂并死亡,却完全无视了它在过去几十亿年的永生!当我们看清楚这一点的时候,我们就可以知道,许多关于细胞衰老和死亡的原因的假说,都是不靠谱的。任何的假说,都必须首先面对这个细胞在过去的几十亿年的永生这个事实,然后再去自圆其说。
前面我们已经说过了,死亡机制,并不是一开始就有的,在死亡机制没有产生之前,许多生物或许是可以“永生”的。并且,我们现在通过比较15岁的人和狗的表现,可以观察到两者巨大差别,虽然他们之间的基因和细胞结构的差别,特别是细胞的内部机制,各种蛋白质的构成其实差别不大。
那么问题又出来了,为什么人和狗会有这样截然不同的表现?答案其实也很简单:由于“永生”是生命的基本属性,在死亡机制产生之后,生命的长短,对于绝大多数多细胞生物来说,其实都是可调的,它是一个可变的变量。某个物种生命的长短,不是内在恒定的,而是自然选择和竞争的结果。当竞争和环境改变了,生命会选择最有利竞争的方向,去改变这个变量。也就是说,复杂生物的生命的长短,不但可控,而且,它们可以通过调整自己生命的长短以适应环境的变化,凡此种种,只是竞争的一种手段而已。而做到这一点,则是通过打开或者关闭死亡衰老机制的开关。生物通过这种方式来进行对自然选择的适应性调节。
为了说明这个问题,我可以再举些例子来说明:
待续............请点击: 死亡起源(八)
备注与参考文献
[14] Carlos López-Otín, Maria A. Blasco, Linda Partridge, Manuel Serrano, Guido Kroemer, The Hallmarks of Aging,cell.2013.05.039
通宝推:lilly,赵沐浴,吃土的蚯蚓,代码ABC,铁手,zen,唵啊吽,北纬42度,河区分,石狼,家园 不同动物不同寿命,也许是导致人类智慧的一个重要前提 生物界里,对于个体来说,最浪费的,可能就是每个个体都要经过从出生到成年的过程,在这个过程中从无到有的累积知识。尤其是对人而言,至少前面的10几年是重复前人所经历过的10几年。要是每个新生能够不但继承父母的基因,还能够自动继承父母的知识和思维,可以想象人类到现在,文明发展程度是什么了。
因为每个个体都要经历从无到有重新掌握知识,所以象狗,寿命10几年,就很难发展出高级智慧,而人能够活几十年,在客观上就提供了高级智慧的时间上的基础。问题是,为什么人活的比狗长。是因为生活习惯、社会结构的变化,在原有基础上挖潜了,还是因为这个人体这个结构本身有一定的优越性?
家园 要这么说,那龟不是要比人类聪明? 或者大象?
家园 铁老大举一反三 呵呵,铁老大举一反三,我们想到一块去了。
关于智慧的发展与寿命的关系,我在死亡起源(十八)——哺乳动物的衰老中有讨论,请赐教。 死亡起源(十八)——哺乳动物的衰老
家园 这段我不是太懂,但觉结论得可能有问题。 “根据细胞学原理,这个细胞,是从几十亿年前,由某个细胞,一直分裂而来的!它穿过了亿万年的时空,经历过可能超过上百亿次的分裂,以“永生”的方式生存到现在,然后才能被我们观察到。”
这段我不是太懂,但觉得结论可能有问题。 举个例子, 某个细胞一变二,二变四。
如果每次分裂可以分清其中某个就是祖先。那四个中有一个生活了两代,一个生活了一代,两个是新生。 但接下来自然选择可能只留下两个新生的。 从而不一定有幸运儿可以代代永生下去。
如果每次分裂不可以分清其中某个就是祖先。那每次分裂都可以看作祖先的死亡,两个细胞都是新生。 如此一来,四个细胞都是新生。 现存的所有细胞也都是新生细胞,不曾生活过亿万年。
当然也可以反过来说每次分裂出来的两个细胞都是永生祖先的克隆。那么所有现存细胞都生活了亿万年。 对于亿万年都没什么代际差异的物种,这么说也可以接受。 但对于代间差异明显的物种,个人觉得这种说法的文学意味大于实际意义了。
望解惑,谢谢。
家园 死亡起源(六)—— 死亡起源 续上: 死亡起源(五)
注:下面这章的内容很重要。经过前面漫长的铺垫以后,很自然的就会往下接着问:为什么?而且随着思考的深入,脑子里面会不停的冒出无数个“为什么?”。关于本章下面的结论,其实在很久以前我就认为结果应该是类似于下面描述的样子,只是当时还缺乏有信服力的证据,而这些证据,却要直到2015年下半年才姗姗来迟.........等了好多年,呵呵。所以,当通过整理这些单独的例子和论文,发现它们之间的联系,并利用这些证据,逐步理顺并揭开了这章所描述的,死亡起源的前因后果的时候,我自己也是很兴奋的。 呵呵。
2.3 “永生”的水螅(Hydra)。
现在我们继续沿着生物演化的轨迹,开始观察另外一种稍微高级一些的原始生物:水螅(Hydra)。作为原始的腔肠动物,他们有两种繁殖方式:出芽生殖和有性生殖。首先,他们可以通过从身体上“出芽”然后脱落,产生一个新的个体。“出芽生殖”是一种无性的生殖方式,通过这种方式,它们可以达到迅速扩大种群的目的。其次,它们也可以通过有性生殖,获得基因以及基因变异的交流和交换,以达到适应自然选择并演化的目的。通过观察他们,我们现在可以看到一些关于衰老和死亡的有趣的现象了。
2015年4月,德国马克斯-普朗克研究所(Max Planck Institute, MPI)在《美国国家科学院院刊》(PNAS)发表了一篇文章[9]。他们的研究发现,在实验室提供的理想的条件下,大部分细胞由干细胞组成的水螅(Hydra)是几乎不会死亡的 。他们观察了2256条生活在清水中的一种水螅超过8年,每天为它们提供理想的条件,结果发现,这些水螅是几乎不会死亡的,而且它们的生殖能力也没有任何下降的迹象。据他们的估算,在人工条件下,即便超过3000年,某些群体至少有5%的水螅依旧可以存活,而且估计这还是因为人工条件不够理想所致。它们如此长的寿命,得益于他们的身体大部分由还未彻底分化的干细胞组成,而干细胞有持续分裂的能力,水螅的身体处在不断更新的状态;水螅的触角和足内的分化细胞在被不断剔除,被从体内“游”来的新细胞代替。而且水螅还具有非凡的再生能力,他们可以在身体大部分缺失的情况下,完全修复自身。如此神奇的“永生”现象,就发生在这么小小的生命体身上。而水螅的这种特性和我们有关系吗? 有的,作为我们演化历史上的祖先,水螅的这种干细胞修复自身的能力和机制,其实就写在我们的DNA里面。
另外,除了个体的“永生”以外,水螅的“出芽生殖”方式 (其实是再生),也意味着水螅个体的死亡的概念变得十分的模糊。因为在某种意义上来说, 出芽生殖方式出生的个体, 我也可以把它看作是和母体对等的一部分,就如同一个细菌分裂成两个细菌,我们无法清楚判断这两个细菌究竟谁是母体一样。
 图19. 2015年4月,德国MPI的研究表明,水螅(Hydra)可能可以“永生”
图19. 2015年4月,德国MPI的研究表明,水螅(Hydra)可能可以“永生” 图19-1. 水螅 (Hydra) 的生命循环。它主要通过“出芽生殖”(Budding)进行大量的繁殖;它也可以通过有性生殖,来产生后代。它们在进行有性生殖时,它的体壁会出现一些“隆起”(Swellings),这些隆起会转变成一个简单形态的卵巢或者睾丸,并产生精子和卵子。精子会被释放到水中和其他个体的卵子受精。水螅(Hydra)和水螅纲(Hydrozoa)的其他物种不一样,它是不会出现水母态的,它终生保留在水螅态 (polyps)
图19-1. 水螅 (Hydra) 的生命循环。它主要通过“出芽生殖”(Budding)进行大量的繁殖;它也可以通过有性生殖,来产生后代。它们在进行有性生殖时,它的体壁会出现一些“隆起”(Swellings),这些隆起会转变成一个简单形态的卵巢或者睾丸,并产生精子和卵子。精子会被释放到水中和其他个体的卵子受精。水螅(Hydra)和水螅纲(Hydrozoa)的其他物种不一样,它是不会出现水母态的,它终生保留在水螅态 (polyps)2.4 多细胞动物程序化死亡的产生和起源。
和水螅同属于近亲的薮枝螅(Obelia),则比水螅更加复杂一些。它主要是以水螅(polyp)的形态存在。它一方面通过无性的出芽生殖产生大量的水螅体以适应竞争所需要的数量。这些大量的水螅,互相连接,如树木般构成树枝状,并通过内部通道互相交换养料。另外,它们还通过所谓的“繁殖水螅”长出雌雄两种水母(medusae),并通过水母进行有性繁殖,这就是所谓的水螅的“世代交替”。
 图20. 薮枝螅(Obelia),它是由很多只薮枝螅个体,搭接而成的。图中的每一个小节,都是一只水螅。它们互相连接,如树木般构成树枝状,并通过内部通道互相交换养料
图20. 薮枝螅(Obelia),它是由很多只薮枝螅个体,搭接而成的。图中的每一个小节,都是一只水螅。它们互相连接,如树木般构成树枝状,并通过内部通道互相交换养料 图21. 薮枝螅(Obelia)的生命循环。它既可以进行无性的出芽生殖,也可以通过水母态产生的精子和卵子进行有性生殖,这就是所谓的水螅的“世代交替”。
图21. 薮枝螅(Obelia)的生命循环。它既可以进行无性的出芽生殖,也可以通过水母态产生的精子和卵子进行有性生殖,这就是所谓的水螅的“世代交替”。薮枝螅的水母型(medusae),生存时间较短,产出性细胞后即死去。它们以水母的形态进行有性繁殖,交换基因,以获得可以适应环境变化的后代。如果把薮枝螅群落看成一棵树的话,它上面的水母,则很像是一朵“花”了。而且,事实上,水母的功能,也和植物的花的功能相同。值得注意的是,近年来的研究告诉我们,植物的花的凋谢和死亡是一种凋亡,是一种器官级别的程序化死亡(Programmed organ Death)[10] 。
和植物的花类似的是,薮枝螅的“花”,也就是繁殖水螅变成的水母,和植物的花一样,开始会死亡(凋亡)了。也就是说,在薮枝螅的“花”上,产生了一个突然死亡的死亡机制!
 图22. 薮枝螅(Obelia)的“花”,也就是它的水母态(medusa)
图22. 薮枝螅(Obelia)的“花”,也就是它的水母态(medusa)而且我们几乎可以肯定,薮枝螅水母繁殖后的迅速死亡,和植物的花一般,也是一种程序化的死亡,是一种自杀行为,是一种器官级别的凋亡(Programmed organ Death)。要知道,妖异的水螅类,很多都具备几乎可以“永生”的能力,而它们的水母如此怪异的突然死亡,很难用正常衰老来解释。我虽然没有找到关于薮枝螅水母的程序化死亡的研究报告,似乎也从没有人做过相关研究,不过,美国西北大学的一个研究团队于2015年7月在《Cell》子刊《molecular cell 》发表了一篇论文 [11][57] , 宣告他们找到了比水螅更高等一些的秀丽隐杆线虫 (C. elegans) 的衰老死亡开关了。
 图23. 2015年7月,美国西北大学的研究者们宣布找到了秀丽隐杆线虫 (C. elegans) 的死亡开关
图23. 2015年7月,美国西北大学的研究者们宣布找到了秀丽隐杆线虫 (C. elegans) 的死亡开关美国西北大学的这项研究是建立在一个十年的研究为基础上的,他们发现了一个基因开关,通过控制这个开关,可以阻止线虫的衰老。这个开关是作用在细胞的热休克反应(Heat Shock Response) 机制上的。热休克反应指的是生物机体在热应激(或其他应激)状态下所表现的以基因表达变化为特征的防御适应反应。它是细胞内部修复机制的一个重要组成部分。热休克蛋(heat shock proteins (HSPs))白戏剧性地上调控是热休克反应的关键部分并且主要由热休克因子引导。热休克蛋白也被称作压力蛋白 (stress-proteins),它通过刺激级联信号通道对冷、热、以及缺氧等进行响应。热休克蛋白通常也存在于那些处于极佳状态的细胞里面。有些热休克蛋白,也叫伴娘蛋白(chaperones), 它们的作用是用来保证细胞的蛋白质在正确的时间,以正确的形状,处于正确的位置。总之,简单说起来,就是一句话,维持正常的热休克反应,对于一个细胞来说,至关重要。美国西北大学的研究者发现,控制这个信号的是线虫的生殖干细胞,保护性热休克反应在线虫性成熟初期的4小时里急剧下降,这也是生殖成熟的精确开始。虽然线虫的行为仍表现正常,但是研究者可以观察到分子变化和蛋白质质量控制的下降。它达到成熟后8小时,所有基因开关都关闭了它们的细胞压力保护机制,从而导致线虫会突然加速衰老和死亡。具体的细节是他们发现对休克反应的抑制是来自于线虫压力基因位H3K27me3标志物的增加,而这又是由通过减少H3K27 脱甲基酶 jmjd-3.1 的表达造成的,这种情况造成的结果是会干扰到热休克因子HSF-1的绑定,抑制对压力反应的正常转录过程。而如果将成熟后的线虫的生殖干细胞去掉,则可以保持jmjd-3.1 的正常表达,从而抑制H3K27me3的累积,维持正常的热休克反应(HSR)。 简单说起来,就是一句话:他们发现了一个重要而且精确的信号开关,如果关闭这个信号的通道,本应该迅速衰老而且死亡的成年细胞依旧可以维持强壮而且健康的状态。另外,和热休克反应相关的基因,在演化上是高度保守的,线虫的这种反应机制,也可以在我们人类身上找到。(注:所谓基因的保守序列(Conserved Sequence ),指的是DNA分子中的一个核苷酸片段或者蛋白质中的氨基酸片段,它们在进化过程中基本保持不变。很多研究者认为,保留序列的基因区域发生突变会导致生命体无法存活或被自然选择所淘汰。)
既然已经找到了线虫的死亡开关,那么这明显是一种全身级别的程序化死亡机制(Programmed organ Death)。虽然没有人研究过薮枝螅水母繁殖后的迅速死亡,但是,美国西北大学的这份重量级的报告,已经为死亡的起源提供了重要的佐证。
 图24. 线虫的死亡开关和信号通道是可以被精确的打开和关闭的。生殖干细胞给出的定时信号会抑制Jmjd-3.1,抑制细胞的热休克蛋白(HSP)的产生,并抑制热休克应激反应(HSR);当去掉生殖干细胞以后,Jmjd-31和热休克蛋白(HSP)以及热休克反应都恢复正常。
图24. 线虫的死亡开关和信号通道是可以被精确的打开和关闭的。生殖干细胞给出的定时信号会抑制Jmjd-3.1,抑制细胞的热休克蛋白(HSP)的产生,并抑制热休克应激反应(HSR);当去掉生殖干细胞以后,Jmjd-31和热休克蛋白(HSP)以及热休克反应都恢复正常。至于为什么薮枝螅要在水母上产生死亡机制,我的观点是:对于薮枝螅来说,它的水母的作用,就是植物的“花”的作用。而薮枝螅的主体,也就是水螅,则是它的主干——它就和一颗树的主干一般。当它的水母完成配子生殖,产生新的交换了DNA的浮浪幼虫后(水螅的幼体),它的任务就完成了,水母作为一朵“花”,它已经没有了存在的必要,所以作为“花”,它触发一个自杀机制,从而自然凋谢(凋亡)是再自然不过的事情了。另外,薮枝螅只有通过幼虫到水螅,才能完成新一轮的DNA交换的循环,这是它们产生水母来实现有性繁殖的目的。当幼虫出生之后,旧水母体内所携带的DNA,已经是老旧的基因了,它作为个体,已经没有存在的必要了,如果它也和水螅一般不死亡,这些水母便会干扰新的DNA的再演化和再循环。所以,在作为水螅的“花”的体内,也就是“水母态”的体内产生这样的如植物的花一般凋谢的机制,会是一个非常正常的自然选择。
总之,我们现在可以观察到,在生物演化的某一刻,在水母身上,产生了死亡机制,而且这个死亡机制,有强烈的证据表明,应该是一种程序化死亡机制,是一种自杀行为。这个死亡机制杀死了携带了旧的DNA水母,完善了新旧DNA的循环和演化更新。或许类似水母的死亡机制可能产生于比薮枝螅更加原始的物种,但是不管怎样,我们在薮枝螅这里观察到了作为“花”的水母的死亡机制的出现。
和上面两种水螅相近的灯塔水母,则走上了另外一条奇特的道路。灯塔水母 (Turritopsis nutricula)在它生命的各个阶段,包括性成熟后,都可以重新回到水螅型(Polyp)状态,所以有人称之为“返老还童”。并且根据研究,它们可能可以无限重复这一过程。有研究声称,这种水母只要不被吃掉或病死,在理论上或许就可以“长生不老”。我认为灯塔水母这样做只是在某些它们认为必要的情况下,通过回到水螅态,进行它们的无性繁殖而已。1995年的时候,有人做过实验 [12] ,发现通过对它们施加以下压力: (A) 饥饿, (B) 突然增加或减少水温(从22℃至17或27℃),(C) 减少盐度(90%海水,10%蒸馏水,S = 33‰), (D) 用剪刀进行机械损伤等的情况下,它们就会被诱导转变为水螅态。另外,2015年12月21日,厦门大学发表于《PLOS One》期刊的一篇文章表明,钵水母纲的海月水母(Aurelia spp.)也可以发生成体生活史逆转,这是在钵水母纲(Scyphozoa)中首次发现这种现象[59]。
灯塔水母和海月水母的这种策略,我个人认为,这只是为了在不同情况下进行不同的繁殖,以此维护种群数量的一个手段罢了,我们不必过于执着和拘泥于它们的“永生”,因为“永生”本就是生命的基本属性,并不值得过分的大惊小怪。生物在竞争条件下为了维护种群数量所采用的策略,经常远比我们想象的更为复杂。比如在某些情况下,一些昆虫,比如某些蝗虫和蚜虫,甚至会如它们的祖先水螅般进行单性的孤雌生殖。它们通过孤雌生殖可以短时间内迅速扩大种群的数量以获得竞争上的优势。具有孤雌生殖能力的昆虫,只需要有一头雌虫传播到某地,就能独立完成繁殖建立种群。而且现在我们知道,许多生物都具有这个能力。这其中包括许多种植物、昆虫、蜥蜴和鱼类。
其实我在这里举出灯塔水母这个例子的主要目的,是要告诉大家,水母类是可以非常任性的。某些水螅以及水母的“水螅态”很可能可以永生,而它们的“水母态”呢,可以演化出“程序化死亡”的自杀机制。也就是说可以死亡了。然后,它的水母态若是突然改变主意了,想“永生”了,就可以和灯塔水母一般,就“永生”了。总之,它们想怎么玩就怎么玩——想自杀就自杀,想“永生”就“永生”,非常的随意而且任性。
 图25. 可以“返老还童”的灯塔水母(Turritopsis nutricula)
图25. 可以“返老还童”的灯塔水母(Turritopsis nutricula) 图26. 2015年12月21日,发表于PLOS One的一篇文章指出,和灯塔水母类似,钵水母纲的海月水母(Aurelia spp.)也可以发生成体生活史逆转
图26. 2015年12月21日,发表于PLOS One的一篇文章指出,和灯塔水母类似,钵水母纲的海月水母(Aurelia spp.)也可以发生成体生活史逆转2.5 由“花”转化成的生命。
如果说薮枝螅(Obelia)是同时采用“水螅态”下带有永生性质的无性繁殖和对于它们的“花”,也就是“水母态”来说,具有“程序化死亡机制”的有性繁殖的话。那么它们的近亲,也是演化得更加高级的钵水母纲(Scyphozoa)的水母们,则在生殖过程中,开始逐步抛弃水螅型和水螅型的无性繁殖,而直接采用水母型的有性繁殖了。这很可能是由于在自然选择中,采用有性繁殖的水母,因为通过交换基因,而获得了更加适应自然选择后代;另一方面,水母拥有更好的游泳技能,也因此拥有更大的自由度,由此在竞争中也获得更加有利的地位。长此以往,本是作为“花”的角色的水母,在演化过程中开始喧宾夺主,脱离了他们的主干,替代了水螅型而独立存在并演化。这些“花”从而成为了这个世界的新主角。
不过也正因为它们曾经是会“凋谢”的“花”,而花是会程序化死亡的,从此开始,程序化死亡机制也就在它们的后代中埋下了。
 图27. 钵水母纲的霞水母(Cyanea capillata),一朵独立演化的“花”。
图27. 钵水母纲的霞水母(Cyanea capillata),一朵独立演化的“花”。另外,虽然我们已经找到了多细胞生物死亡的原因,不过,我非常怀疑这些简单的生物是否已经演化出了衰老机制。因为看起来它们还没有学会如何衰老。它们似乎只是刚刚掌握了一个on/off开关。打开就是“永生”,关闭就是死亡。事实上,观察起来,它们在生殖之前的表现也没有一丝一毫的衰老的迹象,表现出来更多的还是那种妖异的“永生”的模样,甚至灯塔水母在性成熟后,还能够返老还童。同样妖异的还有人体内的血吸虫。同样有着浑身干细胞的它们有着超凡的再生能力,它们甚至可以从一个小组织碎片上重新再生器官[13]。一些研究怀疑它们在死前都不会衰老,而且小小血吸虫的寿命居然可以达几十年之久。另外还有昆虫也是如此,许多昆虫在生殖前(通常也是临死前)都精力充沛,丝毫看不到衰老的迹象,它们的衰老和死亡往往是在很短的时间内发生的。另外,在本文2.4中提到的美国西北大学观察到的线虫,也是突然的进行有开关控制的程序化死亡。由此看来,生物演化一直顺着水螅到血吸虫,线虫直至昆虫,似乎在它们的大部分物种当中,都还没有演化出严格意义上的衰老来。
另外再比较一下水螅、线虫和大多数昆虫,这三类动物和血吸虫的不同。前面三类的大部分,都是典型的生殖后迅速程序化死亡,而血吸虫,则在整个生命中都在生殖,并且同时保持非常健康甚至是接近“永生”的妖异模样。水母中有些是长寿命的,可以存活好几个月,它们也是如此,在死亡之前,几乎每天都在产卵。由此可见,它们的死亡,一定与生殖相关,生殖系统的成熟一定如美国西北大学实验室的线虫所揭示的那样,为死亡开关的打开提供了某种信号。并且,在有需要的情况下,它们也完全可以如血吸虫般,将死亡开关关闭。由此可见,生命对于衰老和死亡的控制,已经到了随心所欲的地步了。
另外,对于美国西北大学的Richard I. Morimoto所说的,认为衰老不是一系列随机缓慢变化的结果,我对他们仅仅保持有限度的支持。他们的观察结果是从线虫这样非常古老原始,并且寿命只有20天左右的生物得到的,这样极端的生命,的确可以消除许多噪声,获得关于死亡的本质是程序化死亡(Programmed organ Death)的证据。但是,关于衰老这个概念,我个人认为,他们的看法或许还不够全面。因为很可能,在线虫这一级别的生命,还没有能够演化出我们通常意义上的衰老机制。拿一个电风扇来做比喻,线虫可能只是刚刚学会了on/off 开关,至于更复杂的多档变速,乃至无级变速,它们可能还没有能够完全掌握。当然,衰老一定和有机整体的程序化死亡(Programmed organ Death)相关,甚至可能是无数这样的死亡的渐进叠加,但是,在哺乳动物身上观察到的真正意义上的衰老过程,相信应该更复杂,应该还会和内分泌和免疫系统相关。具体的相关机制,还需要在更加高等的生物身上去寻找。
钵水母纲的这些以水母形式为主要生活方式的水母,和薮枝螅不同的是,它们的“花”现在成了主要生命形式,所以,花儿们也需要时间去生长、发育、繁殖,于是它们需要将水母阶段的寿命延长以完成生命周期。所以它们的水母阶段的寿命要大大超过薮枝螅水母。有些多次繁殖的水母,寿命可以长达几个月。
总之不管怎样,当生物演化到这个阶段,衰老(非常迅速的衰老)和死亡这两种机制便都产生了。
生物演化树告诉我们,这些水母中的一支,很可能是我们人类的祖先。所以, 现在我们知道了:为什么我们会死亡? 只因为我们本来就是“花”,而花本就会凋谢的。另外呢,在观察到水母的死亡的时候,我们应该同时注意到水母在水螅态的“不死”,因为他们本就是同一种生物。
自此,“程序化死亡”和具有无限自我修复能力的“永生”这样一对非常矛盾的东西,就同时存在于各种多细胞动物体内了。这种“永生”和“死亡”之间的纠葛,会在生物演化过程中,在多细胞动物体内一直纠缠,一直到我们人体内部亦是如此。正是由于这两者之间的共同作用,使得生命获得了具有相当广泛的对环境的适应能力,也使得生命的演化,从此变得更加多姿多彩。
===============================
说个题外话,本章中的几个例子,都是分别花了8年,乃至10年才出的成果。至于灯塔水母,找资料的时候看到一个日本教授,京都大学的水生物学家新久保田,从1976年开始,花了几十年时间研究灯塔水母,试图从这些水母身上身上发现“永生”的奥秘。正是因为这些人的几十年的研究,才有了这个章节的有趣,也令人激动的结论。
待续......... 请点击:死亡起源(七)
备注与参考文献
[9] Ralf Schaible, Alexander Scheuerlein, Maciej J. Danko, Jutta Gampe, Daniel E. Martínez, James W. Vaupel, ?Constant Mortality and Fertility over Age in Hydra“,
[10] HILARY J. ROGERS, Programmed Cell Death in Floral Organs: How and Why do Flowers Die? Ann Bot. 2006 Mar; 97(3): 309–315.
[11] Simple Flip of Genetic Switch Determines Longevity in Animals. http://www.northwestern.edu/newscenter/stories/2015/07/genetic-switch-determines-longevity-in-animals.html
[12] S. Piraino, F. Boero, B. Aeschbach and V. Schmid, Reversing the Life Cycle: Medusae Transforming into Polyps and Cell Transdifferentiation in Turritopsis nutricula (Cnidaria, Hydrozoa),Biol. Bull. June 1, 1996 vol. 190 no. 3 302-312
[13] James J. Collins III,Bo Wang,Bramwell G. Lambrus, Marla E. Tharp,Harini Iyer & Phillip A. Newmark,Adult somatic stem cells in the human parasite Schistosoma mansoni,Nature (2013) doi:10.1038/nature11924
[57] Repression of the Heat Shock Response Is a Programmed Event at the Onset of Reproduction,DOI: http://dx.doi.org/10.1016/j.molcel.2015.06.027
[59] Jinru He , Lianming Zheng , Wenjing Zhang , Yuanshao Lin , Life Cycle Reversal in Aurelia sp.1 (Cnidaria, Scyphozoa),Published: December 21, 2015 DOI: 10.1371/journal.pone.0145314
通宝推:lilly,tiderew,伯威,witten1,代码ABC,铁手,zen,唵啊吽,浣花岛主,gp4444,四条,何故悲秋扇,北纬42度,盲人摸象,老老狐狸,唐家山,富柜,西电鲁丁,河区分,天白,石狼,乔治·奥威尔,joyfm,大山猫,陈王奋起,家园 死亡和开花 以前看到过唵啊吽:死亡冲动与性高潮。思路有很多类似的地方。
谈点个人看法。植物的死亡和开花,我觉得好理解。因为环境恶化,植物面临生存问题时,开花形成种子。种子可以更容易熬过恶劣环境,等到条件合适时,再获得“重生”。这种情形,拿草来说,几乎每年都在重复。春天来了,雨水充足,温度合适,于是种子发芽。等到快到夏天,雨水减少,气温升高,就结了籽,草自己慢慢干枯。所谓“离离原上草,一岁一枯荣”。
动物,原来不是很好理解。动物和植物不一样。小动物出生它一样要吃要喝,不象种子可以埋在地下躲过干旱。现在明白一些,植物的死亡和开花,更象是一种被动的策略。而动物的死亡和“开花”,更象是一种主动的策略。