主题:【原创】死亡起源 The Origin of Death -- az09
家园 这些病毒的释放,应该和人类活动有关。 这些病毒的释放,应该和人类活动有关。其实这些病毒都早已存在了,只是,这些病毒本来,只是被局限在一个局部地区传播。而人类的活动,特别是交通工具的日益发达,给这些病毒的传播,提供了许多的便利。总之,其实还是人类自己折腾的。
至于病毒的寿命,其实病毒这种非常原始东西,不太好讨论寿命。病毒的适应能力本就很强,因为病毒太容易突变了。有些病毒简直就是故意设计成容易突变的。比如,人体的DNA,为了保持稳定性,是双螺旋的两条链的,链之间有非常强的化学键,这可以保持稳定性。而不少病毒,是只有RNA的,而RNA是单链的,稳定性显然要比DNA差很多,不过,它们也因此更加容易突变,或许因此也就提高了适应能力。呵呵。
通宝推:唵啊吽,家园 时间上的传播,到空间上的传播。 病毒和原始宿主互相进化是时间方向上的, 病毒不急着杀死宿主,但又有足够的传染性,让宿主一代代的传染下去。宿主也同时进化免疫系统,解毒病毒,生出健康的下一代。 如果互相进化方向是病毒越来越毒,原始宿主越来越免疫,那难免不断强化的病毒会跨跃物种,跳到会很快被病毒杀死的物种身上。和免疫抗生素的超级细菌出现一样。
家园 埃博拉塞卡爱滋都来自非洲
家园 有个问题要请教,关于永生 生命延续的定义是什么?以文章中所说的大肠杆菌的分裂生殖为例,下一代中包含错误基因的那个当然不是本尊,但正确基因的那个为什么能看成为本尊呢?他们基因100%完全一样么?如果不一样,哪怕非常小,那么称大肠杆菌永生逻辑上说不过去。
其次,假定基因完全一致即可称为本尊,那么假定克隆人完全成功,是不是人类就永生了?那样感觉上似乎有些不对。盼请解惑。
家园 您提了一个很好的问题。 您提了一个很好的问题。
首先解释一下,所谓细胞“永生”的定义,就是指的是细胞的拥有不断分裂的能力。通常指的是细胞不会因为DNA的损伤或者端粒的缩短,导致不能分裂。这个概念是在研究体细胞的时候发展起来的,一开始生物学家以为体细胞是可以无限制分裂的。后来才知道不是这样,有所谓的Hayflick limit (1961年)。现在有不少关于细胞永生的专业术语,比如“细胞的永生化” (cellular immortality)说的也是细胞拥有这种无限增殖能力。
上面的解释,听起来有些玩赖,那么我试着让多细胞的我们,理解单细胞的永生和多细胞的我们有什么关系。比如文章后面提到的“永生"的水螅,以及可以通过营养生殖,达到事实永生的一些植物,它们的永生,是所有的体细胞都不死亡吗?显然不是,它们的永生,是通过拥有”永生“的干细胞,不断的产生新的组织来修复和发展自身,从而达到永生的效果。多细胞生物,首先需要拥有能够“永生”的干细胞,才能够实现永生。 生命一直都是这样,处于一个动态的平衡当中的。
至于讨论的“本尊”,这样的概念,对于被观察的细菌来说,只是我这个观察者给他的定义,我说他是本尊,它就是本尊。对于它自己来说,它每20分钟分裂一次,谁是本尊呢?我们可以说两个都是。而且,即便和细菌本身,是否本尊也和他的DNA是否变化无关,它的DNA即便变化了,它也还是它自己,正如我们身上的不少体细胞的DNA其实一直在突变的,我们也还是我们。“我”这个概念,有时是一个...........哲学概念。对于细菌这种东西,我们不能用多细胞的我们的个体概念来理解。你例子中举的克隆人还不够科幻,我来举一个例子:想象一下,假如你可以如细菌一样分裂,同时还能复制思想和记忆,然后,分裂成两个一模一样的人,而且还有一样的思想和记忆,那么,请问,哪个才是你本尊呢? 呵呵。 头大了吧? 呵呵。而细菌面临的,就是这样的情况。然后你再细想一下,如果你拥有这样无限分裂的能力,那你显然是“永生”的,至于哪个才是你本尊,重要吗?呵呵。每一个复制出来的你,都将有独立的自我意识,都会理所当然的认为自己才是你的“本尊”,而最终决定谁才是你本尊的,其实只是旁边的观察者,而不是你本人了。( 如果还不明白的话,再设想一个场景:在分裂之前,你深爱着一个人,至死不渝的那种,呵呵。在你分裂之后,毫无疑问的,所有的你,都还会继续深爱这个人。但是,她,只能挑选其中的一个“你”,其他的“你”,都只能被淘汰,在一旁捶胸顿足.......。所以,最终决定哪个“你”才是"你"的,是一个旁边的观察者——就是那个她。呵呵)
或者,你可以这样想,把那个细菌,看成一个受精卵,然后,把它分裂的所有细胞,看成一个整体,一个单个的整体。如果这个单个的整体,可以无限增殖而永生的话,那这个细胞就是永生的,如果这个细胞有分裂限制,不能无限制增生,那么它产生的这个细胞群所构成的“单个整体”,也就不是永生的,因为它终会死亡。
家园 死亡起源(九)—— 表观遗传与寿命调节 续上:死亡起源(八)
3.2 衰老与死亡的钥匙。
如前文所述,当生物的自然死亡的真相和真正的主要原因,开始逐渐的指向于它们的死亡开关的适时开启的时候,我们很自然的会开始继续问:那究竟是哪些因素导致了我们死亡开关的适时开启呢?
影响衰老和死亡的因素非常的多,比如前几节中提到的,《Cell》杂志的综述就列举了9大项,另外还有许许多多的因素。这些因素,都毫无疑问的会影响到生物的寿命,而且它们大多都已经有了非常广泛的研究做依据。但是,这些五花八门的各种因素以及相关的研究,也因此构成了一个非常大的噪声和迷雾,它让我们无所适从。并且,这些研究还和各种商业利益联系在了一起,各种相关产品以及广告和宣传,可以说是铺天盖地,很多的宣传都宣称,他们发现了“青春的源泉”,“抗衰老的秘密”,这也同样影响了我们对事物本质的判断。
不过,拨开重重迷雾,从演化的角度上看,我所观察到的与衰老与死亡相关的最重要的钥匙应却该是如下几项:
1. 生殖;2. 压力;3. 遗传与变异 (包括表观遗传) 。
不过在讨论这些因素之前,我们首先需要了解一个有趣的新兴学科: “表观遗传学(epigenetics)”。之所以要简单介绍表观遗传学,是因为,讨论生物死亡和衰老机制,表观遗传是无法绕过的,它参与了演化过程中非常重要的反馈循环。
首先要指出,“表观遗传学”实在是太新了,许多的关于它的机制,尚处在“黑箱”当中。它的真正的发展,也只是最近10年的事情,它的许多细节,还很不清楚,很多的研究,也只是在描述一种观察到的现象,而具体基因的修饰位点却还没有找到。另外,许多的遗传机制,比如,有些可遗传的基因修饰位点,究竟是如何遗传给下一代的,似乎也还不十分清楚。(注:有研究表明,lncRNA参与了表观遗传学修饰的指向目标位点,它可能参与并介导了表观遗传学修饰及其跨代遗传的现象)。
关于表观遗传,每个月还都有大量的新文章面世。我根据我可以找到的资料加上我的理解,写出下面的这一段,基本上是一种相对宏观的描述。过于微观的描述,相信可能超出了绝大多数人的能力,因为这里面的东西太庞杂了,也存在着太多的未了机制,每一项具体的研究,都够一个团队搞几年的了,呵呵。不过,在讨论本文的时候,我们只需要知道宏观层面的结果,暂时也就足够了。因为所有的知识都是可以按抽象程度不同,进行抽象分层的,就如同计算机软件设计和各种协议的分层模型结构(Layer) 一样。大部分的软件设计程序员都无需知道太多的底层原理,他们只需要在自己相关的层面(Layer)上编程与建模,就可以编写出有效的程序。所以,我们在更高的抽象层面讨论,同样也可以得出有趣的结果。而事实上,本文全文,都在基于一个相对宏观的层面上讨论问题,只在需要讨论微观的时候,才举出微观的例子。这是因为,许多微观的研究例子,实际上只是在针尖上做文章,如果我们只是一直在针尖上打转,管中窥豹,我们是看不见事情的全貌的。(根据宏观层面的观察,而得出有趣结论的一个典型例子,便是达尔文和他的《物种起源》了。他在写这本书的时候,甚至连基因是什么都不知道,不过,他在他当时的技术条件所能达到的层面上,进行抽象和总结,同样可以写下他的《物种起源》。)同时,我知道,西西河里,藏龙卧虎,希望有这方面专业的方家可以出来指正。谢谢。
3.2.1 表观遗传学(epigenetics)
达尔文的生命演化论,由于过于强调突变的作用,因此曾经被人诟病为:“在一阵狂风的作用下,将地面上的一大堆零件组装成一架波音747”。那么我们不妨来看看,人类又是如何升级并“演化”一架波音747飞机的。其实和绝大多数工程设计一样,飞机设计人员对产品的升级,一直都是以一种螺旋形反馈递进的。设计人员不断通过分析并解决现有产品的问题,在产品的全寿命期间,对产品在使用过程中出现的问题进行修正和改进。与此同时,飞机的机身以及各个零部件和发动机的生产厂家,会定期发布各种服务通告(Service Bulletin),提供各种改进措施和改良过的零件,并要求在规定的时间内执行和更换,以此保证飞机的飞行安全。同时,设计人员也会根据使用过程中获得的反馈信息,对下一代产品进行改进。
在了解了人类对工业产品的设计和“演化”思路以后,我们再转回头看看生物的演化。在演化论的发展史上,一直有两个不同的学派:达尔文的“自然选择”和由法国生物学家拉马克提出的“用进废退”。在过去的100多年当中,达尔文的“自然选择”获得了压倒性的胜利。而拉马克则一直是作为一个错误的反面教材,被写在教科书里面。有无数的基因和分子层面的研究都告诉我们达尔文是对的。
不过,我认为达尔文的“自然选择”和传统的遗传理论中,一直有一个非常奇怪的问题——为什么生物就不能够将自己生活中采集到的信息,通过遗传的方式遗传下去呢?虽然所有的分子层面的遗传研究都告诉我们,没有发现生物拥有采集身体信息,并可以改写DNA碱基排列顺序的机制。但是,直觉告诉我们,类似的信息采集系统应该是可以演化出来的。因为只有这样,生物的演化才能和达尔文的“自然选择”一起,形成一个完美的反馈闭环,物种才能迅速的演化。仅仅靠基因突变这一种方式演化,客观上来说太慢,太随机,也太不可靠了。特别是对于越来越复杂的物种来说,如果没有足够的种群数量,单单依靠随机突变和自然选择,就好比掷骰子一般纯粹靠运气,这其实是一种不稳定的演化结构,这样的不稳定结构很容易在亿万年的传承过程中或崩溃或被淘汰而消失。即使是放在十万年的时间长度来看,单凭气候的剧烈变迁,就足以让大多数物种消失了,更遑论千万年乃至亿年。
我们再展开讨论一下:假设环境或者竞争的一个突然的短期变化,需要我们的身体的某个基因,在短期内就出现某个特定指向的性状的改变,但是,我们也知道,我们体内的基因,动辄几万个,我们如何保证,我们可以通过“掷骰子”的方式,去精确的指向所需要的改变?首先几万分之一的概率,就已经很小了,并且基因本身又是一串长链,它或许还需要精准的指向某一小段基因片段,而且,基因自己突变的概率更小,两者相乘,则接近0了,如果没有足够的种群数量,加上足够的时间,这基本上是一件不可能完成的任务。那如果再复杂一些,变化的环境需要我们同时出现几个性状的改变呢?然后,再考虑一个更加复杂的情况,即便它运气很好,“掷骰子”中奖,获得了所需要基因的变化,但如果这个短期的环境的的变化消失呢?那又如何变回去?要知道,这种情况其实在生命的经历中非常常见。
所以,以其相信高等复杂生物的演化是通过基因突变累积的,我宁可相信他们主要是通过类似“转座子”(Transposon)的方式,以这样的基因的“剪切与粘贴”或者“复制与粘贴”造成的,至少,这样看起来还靠谱些。
另外,从技术的角度上来讲,可以无中生有,自己逐步演化出来的高度复杂的生命,已经向我们展示出了一套无与伦比的技术。既然已经拥有了这样的技术,那么,再演化出一套具有信息采集并可以遗传反馈的系统,以主动改写的方式加速或者调整自己的演化,以获得更好的竞争地位,在技术上并不应该是一件多么困难的事情。这实际上也是我们人类在工业生产中经常采用的策略。那么这里就有一个非常简单的逻辑推理了:既然在技术上并不困难,那么只要有一种生命率先演化出这样的机制,那么,它就一定会处于竞争顶端的位置。就好比龟兔赛跑一般,准确的说,或许是汽车和乌龟在赛跑。那么,拥有这样机制的物种的后代相对于没有这样机制的物种,就会处于竞争中绝对优势的地位。没有这样机制的物种,没有特殊情况话,会被迅速的淘汰。那么,整个世界,就会处处充满拥有这种机制的物种所产生的后代,以及它的后代演化出来的各种物种了。
而且事实也是如此。我们通过观察地球的生命演化史会发现:如果按照达尔文的演化论以及传统基因遗传和变异的观点,我们很难以此解释诸如“寒武纪生命大爆发”这样的历史事实。寒武纪大爆发,是相对短时期的进化事件,开始于距今5.42亿年前的寒武纪时期,化石记录显示绝大多数的动物“门”都在这一时期出现了。它持续了接下来的大约2千万年-2.5千万年,它导致了大多数现代动物门的发散。因出现大量的较高等生物以及物种多样性,于是,这一情形被形象地称为生命大爆发。这也是显生宙的开始。寒武纪大爆发的事实上也让提出演化论的达尔文非常困惑,他在《物种起源》中写道:“这件事情到现在为止都还没办法解释。所以,或许有些人刚好就可以用这个案例,来驳斥我提出的演化观点”。寒武纪大爆发说明了两个事实:1. 因为曾经长期缺乏寒武纪之前的化石证据,有人据此质疑,生物的各个门好像是突然冒出来的。演化在这个点,不是“树”状结构,而有点象是“草”状的平行结构。不过,最新有一些化石证据,比如“瓮安动物化石群”和“埃迪卡拉动物化石群”等等 ,指出寒武纪之前,也是有化石证据来支持演化论的思想的。2. 寒武纪大爆发显示,生物的演化速度好像非常的快,似乎显示生命经过一段时间的积累以后,在短时间内,突然喷薄而出,这似乎不是用简单的基因突变就可以解释的。
另外,还有一个令人困惑的地方是,我们可以观察到各种各样的生物的貌似主动的自杀行为。看起来是在用自杀来加速演化的进程。刚刚已经讨论了,如果按传统的遗传变异的观点来分析,匆匆忙忙死亡或许并不是一个好的选择。因为基因突变就好像是掷骰子,丢出的骰子可能好,也可能坏,而且生物越复杂,基因突变所产生的相当部分基因都是有破坏性且不利于后代生存的,这样的结构不是一个稳定结构。而且我们知道死亡是有成本的,死亡并重新产生下一代就意味着一切要从头开始,这是需要耗费大量能量并且面临高昂的机会成本的。打一个不恰当的比喻:武侠小说《天龙八部》中的“天山童姥”,她修炼的是“八荒六合唯我独尊功”。修炼这门功夫有一个特点,就是每隔30年需要“返老还童”一次,每循环一次,她的武功便会精进一层。不过,天山童姥虽然武功盖世,但是,当她返老还童的时候,却居然连虚竹这样一个弱不禁风小和尚都打不过,更何况生命所面临的,是弱肉强食的丛林法则呢?在热带丛林中,即便是鳄鱼这样的狠角色,在幼年的时候也往往会沦为水鸟的食物。如果不是死亡可以获得的好处大大高于个体的生存,没有物种会这么傻——即便曾经出现过这么傻的物种,那也是会被自然选择所淘汰的。
所以,问题就来了:为什没有出现这样的信息采集并主动更新的机制呢,还是这种机制其实早已存在亿万年,只是我们没有发现而已呢?或许,它是用另外一种更加不着痕迹的方式存在着?
最新的表观遗传学为我们找到了答案——至少,是揭示了冰山的一角。关于的“表观遗传学”的报道已经非常多了。我只是简单介绍一下。以下摘抄一段表观遗传学的介绍:
表观遗传学(英语:epigenetics)又译为表征遗传学、拟遗传学、表遗传学、外遗传学以及后遗传学,在生物学和特定的遗传学领域,其研究的是在不改变DNA序列的前提下,通过某些机制引起可遗传的基因表达或细胞表现型的变化。表征遗传学是20世纪80年代逐渐兴起的一门学科,是在研究与经典的孟德尔遗传学遗传法则不相符的许多生命现象过程中逐步发展起来的。表征遗传现象包括DNA、RNA干扰、组蛋白修饰等。与经典遗传学以研究基因序列影响生物学功能为核心相比,表征遗传学主要研究这些“表征遗传现象”建立和维持的机制。其研究内容主要包括两类,一类为基因选择性转录表达的调控,有DNA甲基化、基因印记、组蛋白共价修饰和染色质重塑;另一类为基因转录后的调控,包括基因组中非编码RNA、微小RNA、反义RNA、内含子及核糖开关等……。最近20年的研究发现,表观遗传信息通过有序地开启和关闭基因的表达来调控生物体的生长、发育和分化,其中DNA甲基化(5甲级胞嘧啶)是最重要的表观遗传信息之一。
 图38. 表观遗传,外婆的人生经历,也会在你的基因中留下印记。(图中左半部发黄的照片显示的是外婆,右半部的彩色照片则是外孙女,中间的双螺旋结构,则代表DNA)图片来自《Discover》杂志。[60]
图38. 表观遗传,外婆的人生经历,也会在你的基因中留下印记。(图中左半部发黄的照片显示的是外婆,右半部的彩色照片则是外孙女,中间的双螺旋结构,则代表DNA)图片来自《Discover》杂志。[60]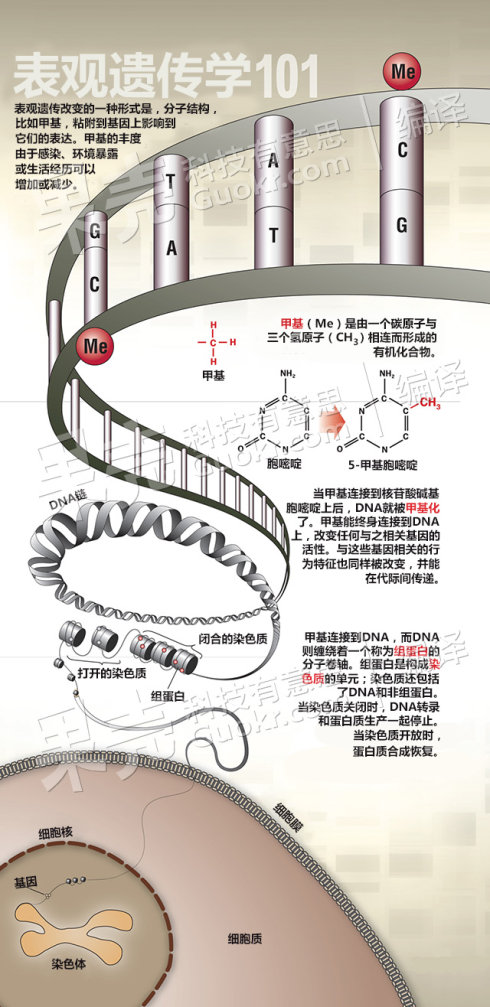 图39. 表观遗传学图解,以DNA的甲基化修饰为例,说明表观遗传原理。图片来自美国《Discover》 杂志,2013年5月文章,由国内guokr网翻译。[60][17]
图39. 表观遗传学图解,以DNA的甲基化修饰为例,说明表观遗传原理。图片来自美国《Discover》 杂志,2013年5月文章,由国内guokr网翻译。[60][17]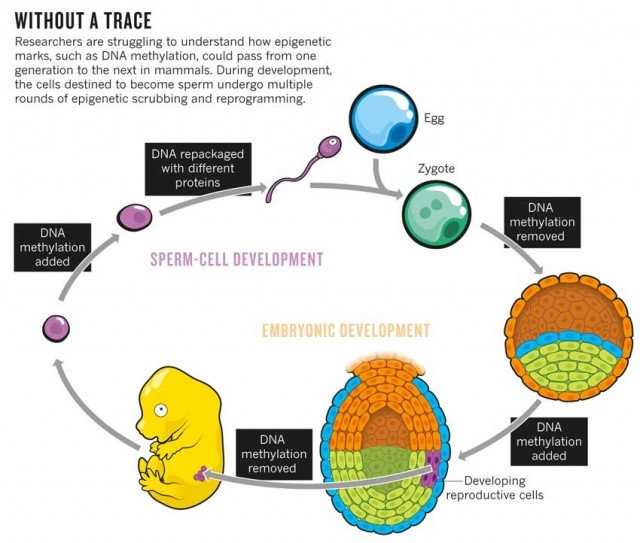 图40. 表观遗传,以哺乳动物的DNA甲基化修饰为例,在它的生命循环和发育过程中,那些要形成精子的生殖细胞,要反复经历多次的甲基化和去甲基化循环(擦洗和重新编程)。而且研究者尚不清楚,表观遗传标记,比如DNA的甲基化,究竟是如何遗传给下一代的。图片来自《Nature》杂志
图40. 表观遗传,以哺乳动物的DNA甲基化修饰为例,在它的生命循环和发育过程中,那些要形成精子的生殖细胞,要反复经历多次的甲基化和去甲基化循环(擦洗和重新编程)。而且研究者尚不清楚,表观遗传标记,比如DNA的甲基化,究竟是如何遗传给下一代的。图片来自《Nature》杂志简单的说起来,表观遗传学就是一个解决了前面所提出来的,生命如何将采集到的信息反馈遗传给后代,以此达到对环境的适应的手段。根据我对表观遗传的理解,我认为,生命采用的方法非常有趣,比我们想象的要更好。现在我们已经知道,我们的DNA中的数据是非常庞大的。但是,有用的基因其实只占整个DNA中非常小的一部分,其中的外显子(Exon),也就是能够制造蛋白质的编码序列,只占总长度的1.5%。大部分的数据就好像是物理学中的“暗物质”一般,无声无息。关于这个现象,一般认为这可能是历史的沉淀,不过对此科学界还存在各种争议,有研究认为这些部分也是有功能的,因为有发现如果敲掉一部分所谓的没有用的DNA部分,会出现许多问题。我们现在随意在我们身上取下一个细胞,只要稍微思考一下,就会发现,这个细胞是通过“永生”的方式,穿越了亿万年的时光,经历亿万次的不断分裂存活下来的。也就是说,我们身上的每一个细胞,它的历史之长,经历之复杂,是我们人类无法想象的。而且关键是,既然它可以历经亿万年存活下来,那么它身上就有一个我们可能没有注意到的,一个其实非常显而易见的事实:我们的细胞的DNA里面可能含有全套的,可以适应这亿万年各种情况的应对机制、措施和处理方案,有一个非常庞大的数据库,这也是它亿万年演化过程的积累和总结。 我们只要稍微回顾一下地球过去几十亿年的历史就可以知道,它身上的这套数据库意味着什么。这意味着我们的DNA在过去的几十亿年里面,已经积攒下了非常庞大的各种应对功能模块!现在,表观遗传学告诉我们,我们DNA里面的这些功能模块是可以根据需要搭建的。我们的细胞可以根据需要,通过开关,把需要的功能模块打开,或者把暂时不需要的模块关闭。比如,我们的多能干细胞可以转化成任何一种体细胞,当它分化为某一个体细胞的时候,可以把某些开关打开或者关闭,于是它就变成了某一个器官的功能细胞了。
而且,最关键的是,现代的研究发现,这些开关,也就是所谓的DNA的甲基化等等,(1)是可以后天获得的。后天的经历,会改变DNA的甲基化等等水平,也就是说,会改变这些开关。(2) 这些后天获得DNA的开关,是可以遗传的! ——至少是可以部分遗传的。这就开始变得有趣了。
我们前面已经讨论过了,如果可以将后天获得的,适应环境变化的一些改变遗传下去的话,那么,这个生物对自然选择的适应性将比传统遗传学(genetic)的单纯靠突变而获得的概率大大增强几个数量级以上。
根据表观遗传学的研究,我们可以看出,生物可能首先是在亿万年的岁月中,通过各种演化和遗传变异,逐步构建了许多基本功能模块。然后,它们可以根据自己的演化和遗传,以及后天的生活经验,选择在细胞和组织分化的时候,或者是在后天的生活过程当中,把自己身上的许多这样的模块的开关去主动打开或者关闭,有选择的表达或者不表达这些模块的功能,以此对环境进行适应。打一个不恰当的比喻,这就好象搭“乐高积木”一样。而且考虑到人类的DNA中,其中的外显子,也就是能够制造蛋白质的编码序列,只占总长度的1.5%,而且蛋白质又可以通过改变空间构象获得更多的变化,我们的DNA很可能只是拿了一大盒积木里面很小的一部分就把我们给搭建出来了。然后再将这些先天或者后天获得的,调整过的,适应环境变化的,许许多多的开关的打开或者关闭的编码和顺序,就象一个被精心调制过的音乐的曲谱一样,以DNA甲基化等等形式遗传下去。于是,生物演化的轨迹,从此开始变得容易而且缤纷多彩了。
生物的这种策略这是非常非常聪明的做法,这比我前面提到的直接改写DNA的碱基排序还要高明。生命通过构建基本DNA模块,使得各种功能模块化,然后再以这些模块为基础,有选择的利用开关去选择和搭建自己的生命形式,并能够在受精后对来自父体和母体的基因进行“重新编程”(通过去甲基化和重新甲基化等),将先天以及后天的经历遗传下去,这才是适应变化的环境以及各种竞争的最科学,最稳定,最高效率的做法。这样做,一方面是不会因为直接改写复杂的DNA碱基的排列而破坏已经技术成熟并且有效的基因和功能模块,同时又通过各种开关来选择打开或者关闭这些模块,获得了应该有的便利性,因此这是一个非常稳定而且高效的结构,一个稳定到可以延续亿万年的结构!真是令人难以想象,这样巧妙的构思,居然是演化自然形成的。
另外,表观遗传还受到不少质疑,质疑之一,便是表观遗传的不稳定性。目前观察到的表观遗传,它通常只能遗传几代,当压力消失后,它并不能稳定的长久的遗传下去。后面我会提到,表观遗传的这种不稳定性,或许正是生命需要的呢。
总之,当我们了解了表观遗传学以后,我们开始知道,生命的演化,的确是拥有一个完美的,包含对后天生活的信息采集、反馈并遗传,以此适应自然选择和竞争的一个反馈闭环的。
那么,我们讨论了这么久的表观遗传以及表观遗传的可遗传性,究竟和我们讨论的衰老与死亡有啥关系呢?答案是关系很大。表观遗传学的突破,让我们有了一个合乎逻辑的衰老和死亡机制的反馈循环。这个话题我们随后会展开。
现在,我们暂时将表观遗传学按下不表,把话题拉回到我们要讨论的衰老和死亡。我们先讨论生殖对它的影响。
待续........ 请点击:死亡起源(十)
备注与参考文献
[17]《祖先的经历,也能遗传给你?》 http://www.guokr.com/article/437295/
[60] Discover, Grandma's Experiences Leave a Mark on Your Genes, http://discovermagazine.com/2013/may/13-grandmas-experiences-leave-epigenetic-mark-on-your-genes
通宝推:昆吾割玉刀,lilly,吴用,tiderew,铁手,唵啊吽,弦音醉舸,浣花岛主,河区分,p47109,天白,加东,富柜,游牧天下,北纬42度,大山猫,积吉,秋哥,石狼,pattern,联储主席,唐家山,家园 DNA里面可能含有全套的,可以适应这亿万年各种情况的应对 这个设计用工程观点看,就是编译开关嘛,程序员也会在代码里加一对编译开关:linux下激活哪一段,windows下激活哪一段
家园 【商榷】表观遗传 这是一个非常有趣的话题。
表观遗传到底是对环境的被动反应还是生物体的主动行为?
也许可以拿人体的免疫系统来对照。(有点忘记当年学过的免疫学了,请达人补充或者纠正)。免疫系统需要面对的外来侵入对象,可以说是数量繁多而且不可预测,但是面对新的威胁,总是能够产生新的抗体,而且这种免疫功能有“记忆能力“。这也是打疫苗获得免疫能力的根源所在。非常有趣的是,免疫系统怎么产生几乎是无穷数量的抗体来应对几乎无穷的抗原?靠的是对已有有限种类的原材料的组装(不记得是基因的组合表达不同蛋白质,还是基因分别表达最后组合成为抗体)。通俗点说,就好像用26个字母组合出单词、短语、句子等等。
我觉得表观遗传是不是和免疫系统类似,因为外界诱发环境的存在,所以原来不表达的基因得以表达。看起来就象是遗传了一样。
如果是能够主动选择性的表达,就象人工智能一样,那就太有意思了。虽然觉得很难理解,但是,既然我们能闭上眼睛思考,这种主动能力应该还是存在,就是不知道怎么实现。
通宝推:az09,家园 美科学家合成出迄今最简单生命形式 英国《金融时报》科学主编 克莱夫?库克森 报道
美国科学家将生命的多余部分剔除,只留下最基本的要素,由此创造出一种合成微生物,这种合成微生物拥有生长和繁殖所需的最低限度的基因信息。
由基因学先驱克雷格?文特尔(Craig Venter)领导的研究人员创造了名为Syn3.0的“最小化合成细菌细胞”,这是他们在2010年创造的曾得到广泛宣传的Syn1.0的后续成果。Syn1.0是首个拥有用实验室化学品从零合成出的脱氧核糖核酸(DNA)的活细胞。
研究人员希望,Syn3.0或其后续样品能提供一个平台,供合成生物学家加入有特定用途的基因,比如生产药品或生物燃料的基因,尽管Syn3.0更直接的目标是更好地理解生命的基本生化机理。
这个项目的研究成果发表在《科学》(Science)期刊上。文特尔博士表示,该项目持续的时间比预期长了四年,揭示出生物学知识中存在“令人吃惊的”空白。
这个研究团队最初的思路是利用科学文献中提供的所有信息,设计一种最小化的细菌基因组,但这条路没有走通。文特尔博士说,这次失败证明“我们目前的生物学知识,不足以让我们坐下来设计一个活的有机体并将它造出来”。
后来,该团队调整了思路,转而研发基于丝状支原体的Syn3.0。该团队展开了漫长的探索,通过逐个剔除再观察结果的办法,观察丝状支原体的901个基因中有哪些是必不可少的。丝状支原体是一种天然的细菌。
就这样,不必要的基因被一个接一个地剔除,最终得到了473个复制和生长所必需的基因。该团队的研究工作是在美国加州约翰?克雷格?文特尔研究所(J. Craig Venter Institute)及其附属公司合成基因组学(Synthetic Genomics)展开的。
编码组成这473个基因的DNA,相当于53.1万个基因代码化学“字母”。这些DNA随后在实验室中被合成出来,合成出的基因组被植入另一种细菌山羊支原体(M capricolum)的壳中,该支原体自身的DNA已被移除。
该合成基因组接管了宿主细胞的生物学运作,产生了一种强健的细菌,该细菌经实验室培养可迅速繁殖,菌落规模每三小时翻一番
家园 换句话说只合成了DNA,没有合成蛋白质
家园 这一段不是很明确 如果我理解的正确,作者是不是说过去生物进化所积累的对所有所经历情况的准备,都已经储备在基因里,在合适的时候打开(也就是甲基化)就可以显现了?这大概是太阳底下没有新鲜事在基因层面的反应吧。不过本人对这个结论存疑 -- 这个观点主要是为了平衡动物的极端复杂性,和基因突变的偶然性和偶发性,不过这个解决方案可能过于昂贵。
- 复 这一段不是很明确
家园 的确值得存疑 的确值得存疑,这个方案的确挺昂贵的, 所以我用了“可能”两个字。
但是,再细想想,有更廉价的方案吗?比如单纯靠基因突变?
我在文章里面写了下面一段,我们思考一下,如何不用昂贵的方案,采用“廉价”的方案去处理?
我们再展开讨论一下:假设环境或者竞争的一个突然的短期变化,需要我们的身体的某个基因,在短期内就出现某个特定指向的性状的改变,但是,我们也知道,我们体内的基因,动辄几万个,我们如何保证,我们可以通过“掷骰子”的方式,去精确的指向所需要的改变?首先几万分之一的概率,就已经很小了,并且基因本身又是一串长链,它或许还需要精准的指向某一小段基因片段,而且,基因自己突变的概率更小,两者相乘,则接近0了,如果没有足够的种群数量,加上足够的时间,这基本上是一件不可能完成的任务。那如果再复杂一些,变化的环境需要我们同时出现几个性状的改变呢?然后,再考虑一个更加复杂的情况,即便它运气很好,“掷骰子”中奖,获得了所需要基因的变化,但如果这个短期的环境的的变化消失呢?那又如何变回去?要知道,这种情况其实在生命的经历中非常常见。所以,以其相信高等复杂生物的演化是通过基因突变累积的,我宁可相信他们主要是通过类似“转座子”(Transposon)的方式,以这样的基因的“剪切与粘贴”或者“复制与粘贴”造成的,至少,这样看起来还靠谱些。
看起来用廉价的方案很难做到,不是吗?
那我们再看事实:事实上,我们体内有许多基因都是演化上保守的,也就是,在演化过程中,基本不变的,要维持这种不变,本身就需要一个相对‘昂贵“的成本。
另外,还有一个更加诡异的事实,是在所谓的”非编码DNA“,也就是所谓的垃圾DNA中,也有许多保守序列,这就更奇怪了。毕竟,我们的”有用“的DNA只占总数的1.5%而已,在非编码区维持这些保守序列,是很奇怪,也是成本很高的事情。
那么,事实上,有没有冗余呢?我认为是有的。比如,我们经常可以看见各种各种样的返祖现象,比如毛孩,我们其实在必要的情况下,还是可以把有长毛发的基因打开的,只是我们不需要了而已。这个基因并没有消失,还藏在我们体内,这就是一种冗余数据的表现。相信这样的冗余数据,还有许多。
还有,我们每一个体细胞,DNA都是一样的,但是,分化后的体细胞,功能各不相同,这意味着这些体细胞,在“数据库”上有相当大的冗余的,再加上它还需要一定的适应性,则需要更大规模的冗余...........
总之,生命似乎为了追求更大的稳定性和适应性,采用了一个大冗余的做法。虽然昂贵,但是却似乎很值得。
这是我的看法。呵呵。
- 复 的确值得存疑
家园 不知道有没有各个物种DNA数量的统计对比 就不说基因的数量吧,这个很难确定,因为有很多象你说的,就是一些看起来无用的DNA片段。就是比较一下碱基对的总体数量。
那些“无用”的DNA片段也是奇怪的事,有没有可能是在繁衍过程中杂交积累的?
家园 这些DNA片段应该是有用的。 这些DNA片段应该是有用的。因为有研究表明,敲掉它们后,会出问题。
我其实有些怀疑转座子Transposon会不会从这些所谓的垃圾片段里面截取一些DNA片段,拿来拷贝或剪切,用来做拼接用啊,或许可以生成一些新的基因。 这或许是一个基因片段库也未可知。我挺喜欢转座子这个东西的,简直就是一个新基因生成器嘛。
也有可能是如你所说的,是“杂交”累积。只是这种“杂交”跨度有多大,似乎还待考。不是说,我们有些基因甚至是由病毒插入的吗?甚至,线粒体也是共生所产生的吗? 呵呵。 我也不知道,实在是有太多未了机制了。
家园 这个DNA甲基化不就是获得性遗传的内在机理? 拉马克包括米丘林的进化理论的被大批特批,但是这是非常诱人的想法。 尽管最近这些年的研究也在非常少的地方证明了获得性遗传,但现代基因理论体系里,对获得性遗传不置可否,认为从来不是是生物进化的主要方式,这个DNA甲基化从分子生物学角度可以更深层次的对拉马克当时提出的总结性理论重新理解。